的irf7和IRF3靶基因的表达受到抑制。在补体和凝结级联途径中,在结合配体(mbl)和三种补体成分(c4、c6和c7)的基因中观察到一致的表达水平升高(图1C)。在抗原处理和呈递途径(图1D)中,逆转录-qPCR阵列数据显示hspa4、hspa5、cd4、calr和b2m中表达水平升高,但是观察到两种主要组织相容性复合体(MHC)即mhc1和mhc2的下调。此外,组织蛋白酶B和L (ctsb和ctsl)以及溶酶体/吞噬作用途径中的必需膜糖蛋白(lamp2)持续上调(图1E)。在白细胞跨内皮迁移途径中,三种重要的整合素(itgb1、itgb7和itga4)和粘附配体(cxcl12)以及关键胶原酶(mmp9)被显著刺激(图1F)。
2. 转基因肾脏免疫相关途径差异表达基因的分析
表2 肾脏中免疫相关途径的DEGs表达水平的结果
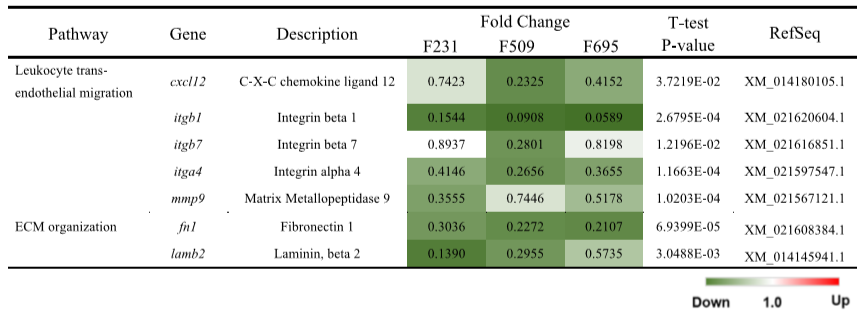
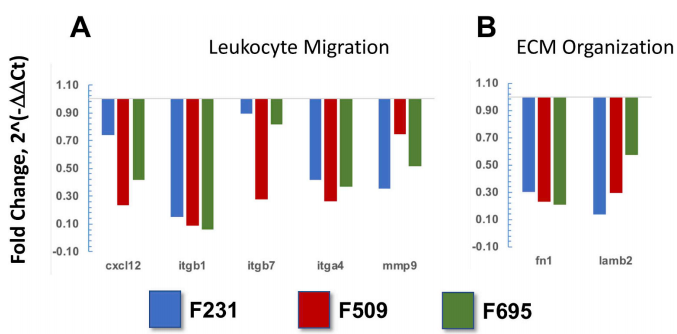
图2虽然白细胞跨内皮迁移途径中关键整合素基因(itgb1、itgb7和itga4)和结合配体基因(cxcl12)的基因表达水平显著升高(图2A),胶原酶基因(mmp9)也上调,但在三种鱼转基因肾脏中观察到调节细胞外基质组织的纤连蛋白(fn1)和层粘连蛋白(lambin 2)基因表达水平的抑制(图2B)。
3. 转基因肝脏免疫相关途径差异表达基因的分析
表3转基因虹鳟肝脏中的免疫途径相关的DEG的芯片分析结果
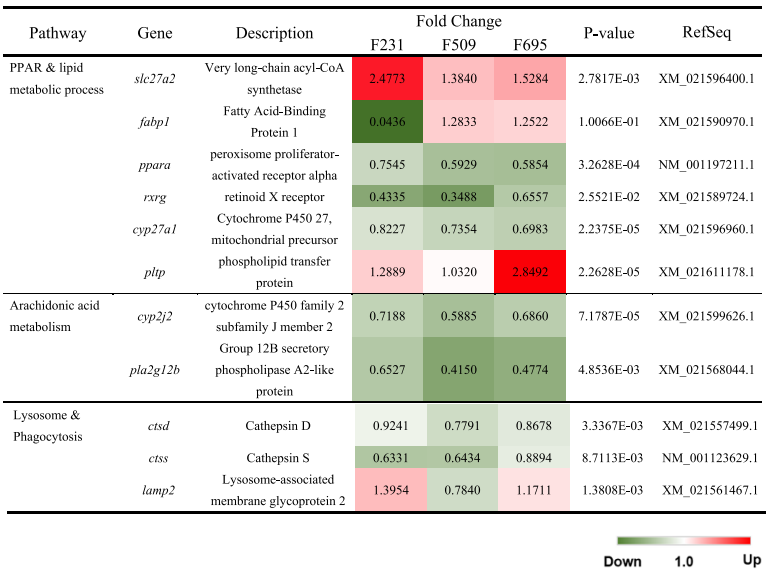
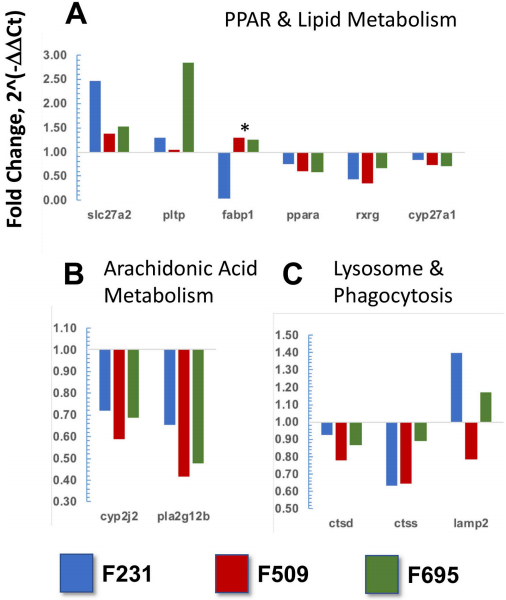
图3 如图3A所示,在三个转基因鱼科中,PPAR信号传导和脂质代谢过程途径中重要的膜转运蛋白/同工酶(slc27a2)和磷脂转移蛋白(pltp)基因的表达差异性增强。相反,转基因肝脏中信号受体(ppar-α和rxr-γ)及其下游靶基因(cyp27a1)的mRNA水平受到抑制。有趣的是,虽然脂肪酸结合蛋白(fabp1)的mRNA水平在F509和F695的转基因鱼科中上调,但它在F231家族中被下调。与花生四烯酸代谢途径相关的DEG被下调,包括cyp2j2和pla2g12b(图3B)。最后,在所有三个转基因家族中,溶酶体/吞噬作用途径中的组织蛋白酶D和S(ctsd和ctss)被一致地抑制(图3C)。有趣的是,膜糖蛋白基因(lamp2)的表达在转基因鱼科F231和F695中得到增强,但它在家族


